|
|---|
|
Предполагается, что расположение и регуляция генов в архитектуре ядра влияют на экспрессию генов. В этой статье показано, что обонятельные нейроны мыши, молчащие гены обонятельных рецепторов разных хромосом объединяются в несколько гетерохроматиновых фокусов. Эти фокусы эксклюзивны для обонятельных рецепторов и образуются в клеточно-специфичной и дифференцировка зависимой манере. Агрегация генов обонятельных рецепторов во время развития синхронизирована с понижающей регуляцией рецептора ламина b (LBR) и может быть отменена эктопической экспрессией LBR в зрелых обонятельных нейронах. LBR вызванная реорганизация архитектуры ядра и нарушение агрегатов обонятельных рецепторов нарушает сингулярность (одиночность) транскрипции обонятельных рецепторов и повреждает целевую специфичность обонятельных нейронов. Наблюдения авторов предполагают пространственное секвестрирование гетерохроматинизированного семейства обонятельных рецепторов в качестве основы для моногенной и моноаллельной экспрессии генов. Пространственная компартментализация генов в ядрах клеток млекопитающих служит регуляторным целям, как предполагают Фразер и Бикмор (Fraser and Bickmore, 2007). Ранее терминами гетерохроматин и эухроматин описывали молчащие и активные участки генома и лишь позже они были охарактеризованы биохимически (Zacharias, 1995). В большинстве типов клеток взаимодействие с ядерной мембраной (пластиной? Lamina) локализуют гетерохроматин по периферии ядра, а эухроматин занимает центральную часть ядра. (Peric-Hupkes и van Steensel, 2010). Микроскопия ядра под большим увеличением выявляет дополнительные уровни организации и компартментализации. Например, транскрипция может быть ограничена специализированными участками ядра или «фабриками» транскрипции, где гены взаиморасположены в определенном порядке (Eskiw et al ., 2010). В итоге межгенные и внутригенные взаимодействия на больших расстояниях внутри генома образуют регулирующие сети, которые контролируют экспрессию генов и дифференцировку (de Wit и de Laat, 2012; Liu et al., 2011; Montavon et al., 2011).
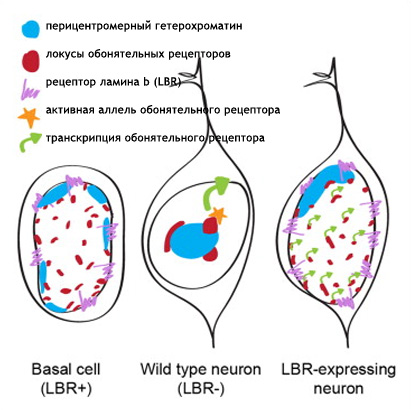
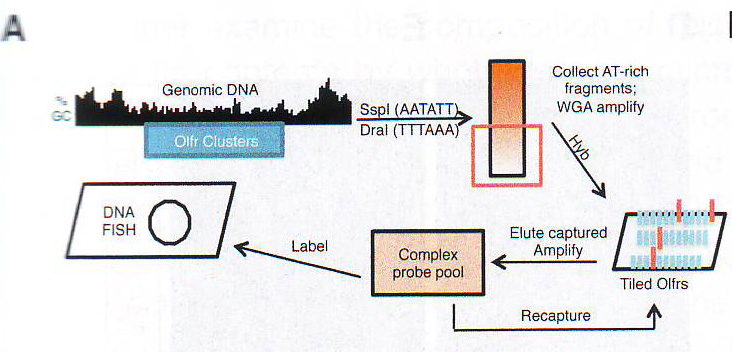
Рис. 1А. Необратимые решения во время развития, принимаемые, например, дифференцирующими нейронами, служат различным эпигенетическим механизмам определять и фиксировать транскрипционный статус клетки на протяжении жизни. Помещая гены в субъядерные отсеки (компартменты), совместимые или не совместимые с транскрипцией, можно финализировать эти решения. Дифференцировка обонятельных сенсорных нейронов – очень яркий пример такого развития. Обонятельные сенсорные нейроны выбирают один из 2800 аллелей обонятельных рецепторов и затем устанавливают стабильную программу транскрипции, которая обеспечивает то, что аксоны этих нейронов образуют специфические гломерулы (Buck и Axel, 1991; Imai et al., 2010). Моноаллельная природа экспрессии обонятельных рецепторов (Chess et al., 1994) вместе с наблюдением, что промотеры обонятельных рецепторов чрезвычайно гомогенны и имеют сходные регуляторные элементы (Clowney et al., 2011) показывают, что одной лишь последовательности ДНК недостаточно для того, чтобы инструктировать экспрессию только одного аллеля в каждом нейроне и что в этом случае налицо эпигенетический механизм. Конечно, открытие гетерохроматинизации вступает в спор с эпигенетическим неопределенным контролем выбора обонятельными рецептоами (Magklara et al., 2011). Так как активные аллели ОР имеют разные модификации хроматина от неактивных ОР и ассоциированных с цис и транс с h усилителем транскрипции (Lomvardas et al., 2006) эта эпигенетическая регуляция может иметь пространственный компонент. Хоть делеция h усилителя транскрипции не имеет значительного влияния на транскрипцию большинства обонятельных рецепторов (Khan, 2011) его ассоциация с активными аллелями обонятельных рецепторов может отражать физическое разделение активных аллелей обонятельных рецепторов от молчащих генов обонятельных рецепторов и их перенос в активирующую ядерную "фабрику". В этой работе авторы изучают значимость организации ядра в экспрессии обонятельных рецепторов. Используя комплексный ДНК FISH образец, который распознает большинство локусов обонятельных рецепторов продемонстрирована OSN специфичность и зависимую дифференцировку от интра и межхромосомных взаимодействий молчащих генов обонятельных рецепторов. Тогда как эти ОР специфичные фокусы колокализованы с метилированными гистонами НЗК9mеЗ, H4K20me3 и гетерохроматиновым белком 1 бета активные ОР аллели имеют минимальное перекрывание с гетерохроматиновыми маркерами и остаются в эухроматиновых областях, предполагая наличие репрессирующего и активирующего ядерного отсека для аллелей ОР. Критична для этой ядерной организации понижающая регуляция и удаление рецептора ламина b из ядерной оболочки OSN. Делеция LBR вызывает эктопическую агрегацию локусов ОР в базальных и поддерживающих клетках основного обонятельного эпителия. Тогда как экспрессия LBR в OSN нарушает формирование локусов ОР, что приводит к декомпактизации гетерохроматина ОР, совместной экспрессии большого количества ОР, в итоге к снижению транскрипции ОР и нарушению OSN таргетинга. Авторы предполагают инструктирующую роль архитектуры ядра в моногенной экспрессии обонятельных рецепторов. РезультатыЧтобы получить картину распространения локусов ОР в OSN авторы создали ДНК FISH образец, который позволит определять независимо расположение большинства локусов ОР.
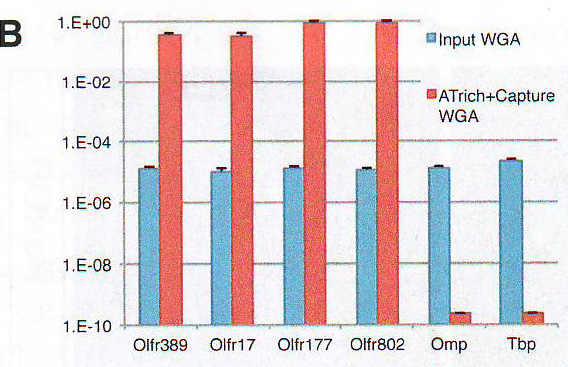
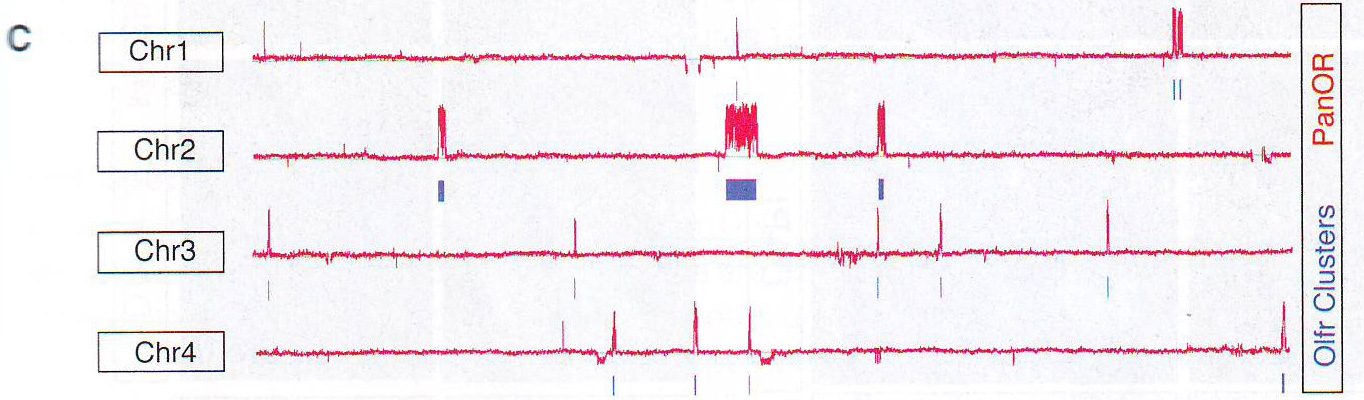
Рис 1B. Для дальнейшего исследования состава этой ДНК библиотеки авторы анализировали ее содержимое путем полногеномной гибридизации эрреем с использованием кафельного (? tiling) эррея, покрывающие хромосомы мыши с 1 по 4. Этот анализ демонстрирует чувствительность и селективность. Cтратегии очистки: образец определяет 340 из 346 генов обонятельных рецепторов (ОР), расположенных на этих хромосомах, 40 из примерно 80 не-ОР генов, расположенных c / s относительно ОР-кластеров (и включенных в захватывающий эррей) и 6 из приблизительно 5000 не ОР-генов (FDR < 0.05, 98.2% чувствительность, 98.4% OR кластерная специфичность, p < 10~ 72 ) (Рис. 1С и Рис. S1B - S1D и таблица S1 доступная онлайн).
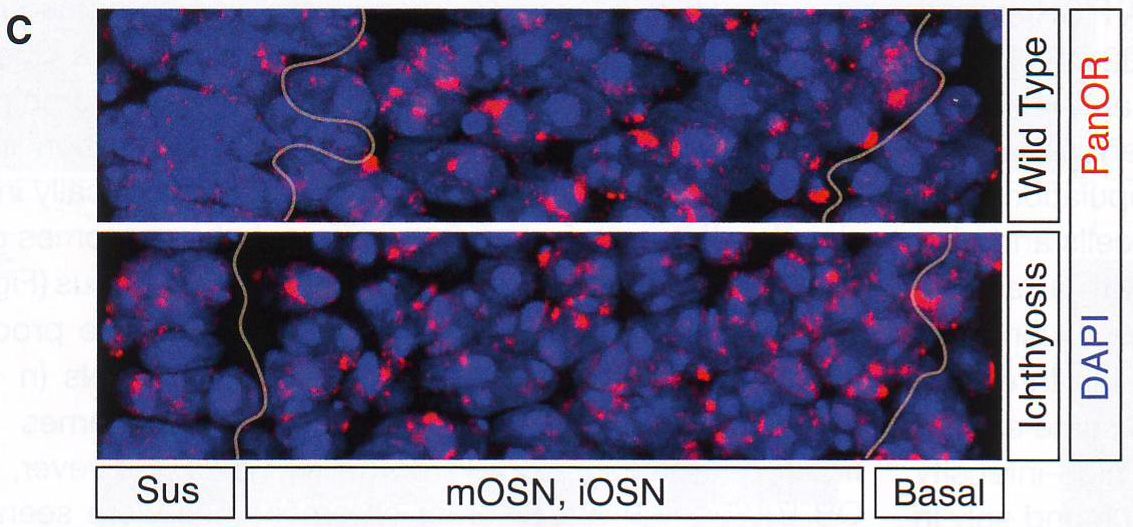
Рис 1C.
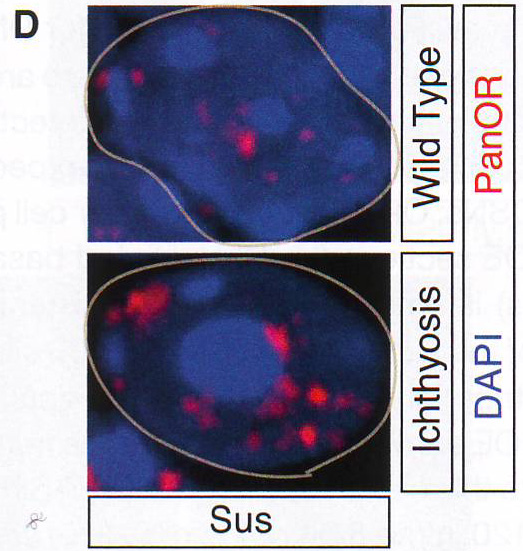
Рис 1D. OSN специфическая агрегация генов обонятельных рецепторовАвторы использовали панОР-библиотеку как образец для ДНК FISH экспериментов на секциях MOE. Хотя существует 92 OR кластера в диплоидном ядре, полный ОР образец определяет в среднем около 5 больших фокусов в OSN (рис 1D). Это неожиданное распределение специфично для OSN: ОР распределение в других клеточных популяциях, представленных в отделах основного обонятельного эпителия (MOE - main olfactory epithelium) недифференцированных базальных клетках и поддерживающих клетках диффузно и более состоит с рандомным распределением 92 ОР кластеров или примерно 2800 аллелей. Подсчет распределения ДНК FISH сигнала в трех типах клеток MOE внутри одних секций в этих же экспериментах поддерживает этот вывод (Рис. S1E-S1F).
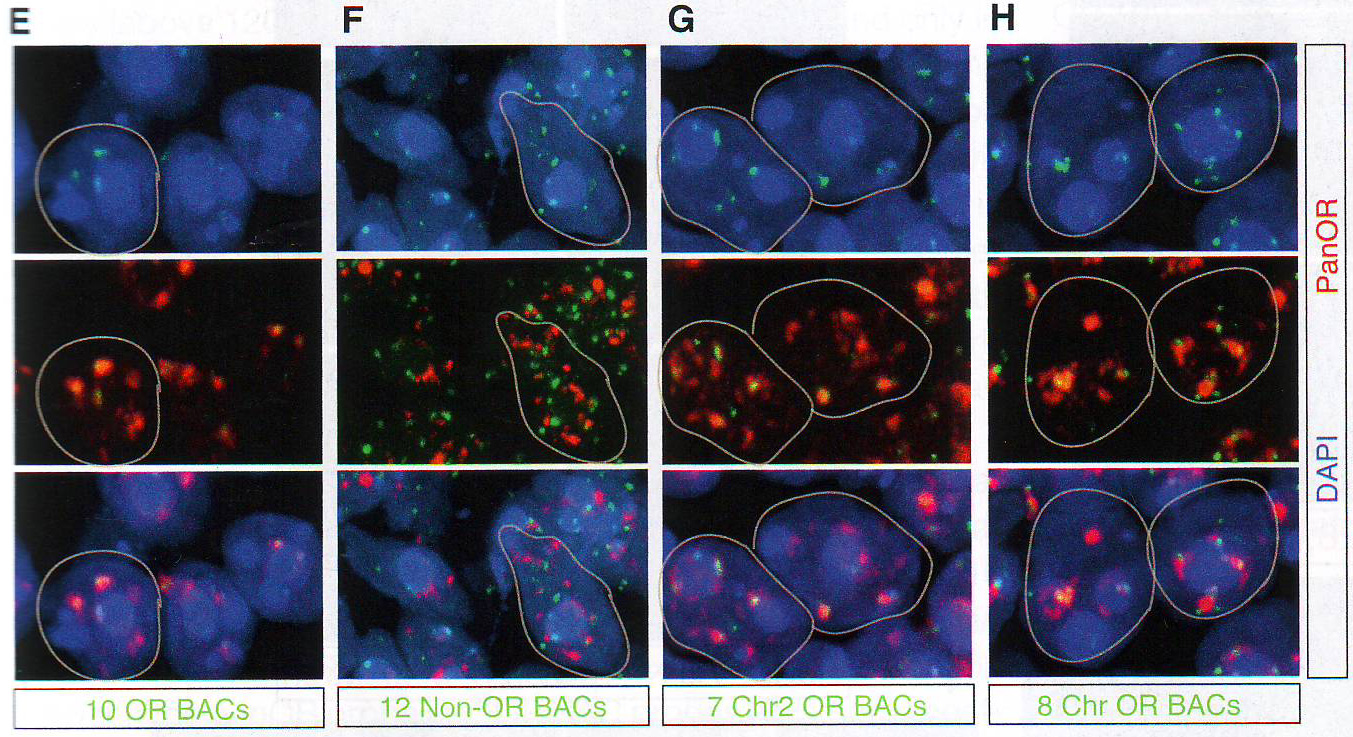
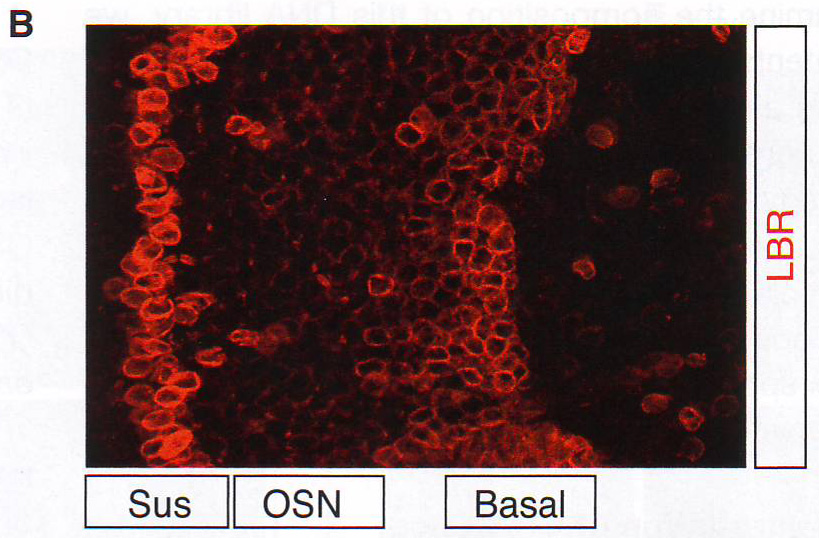
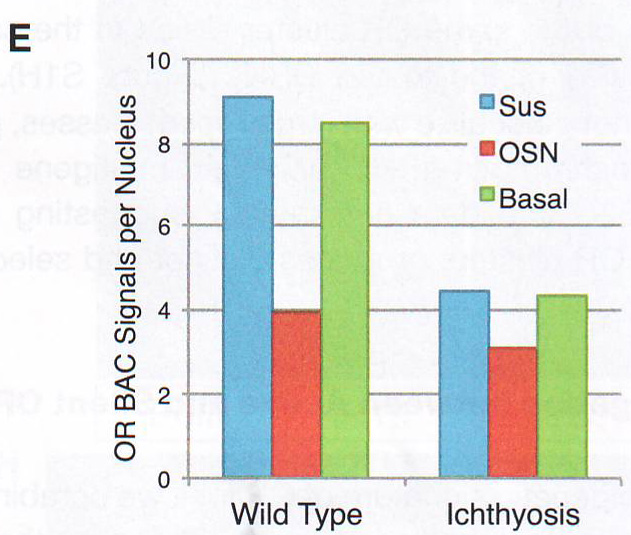
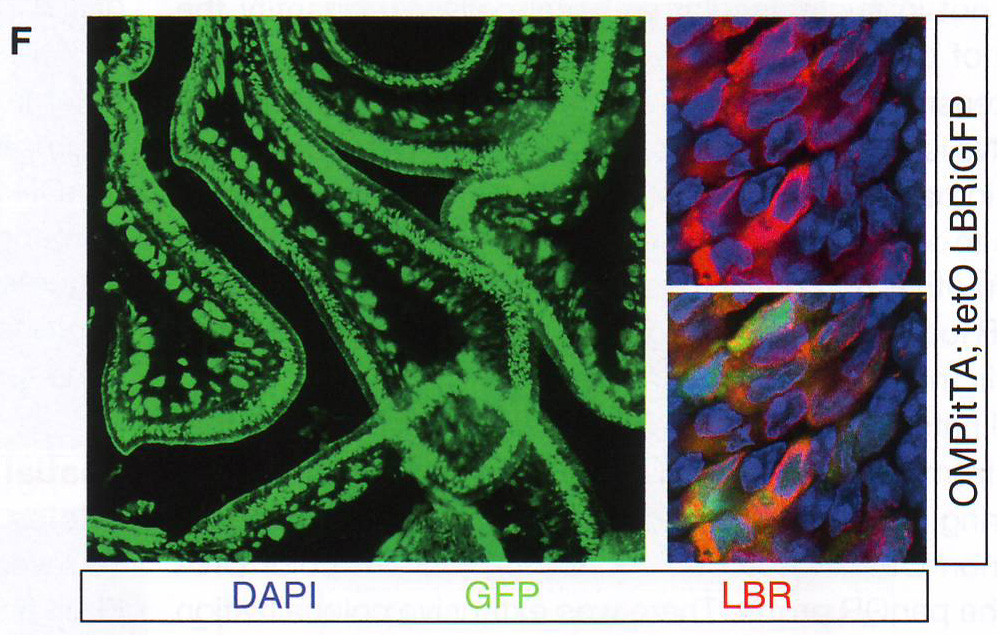
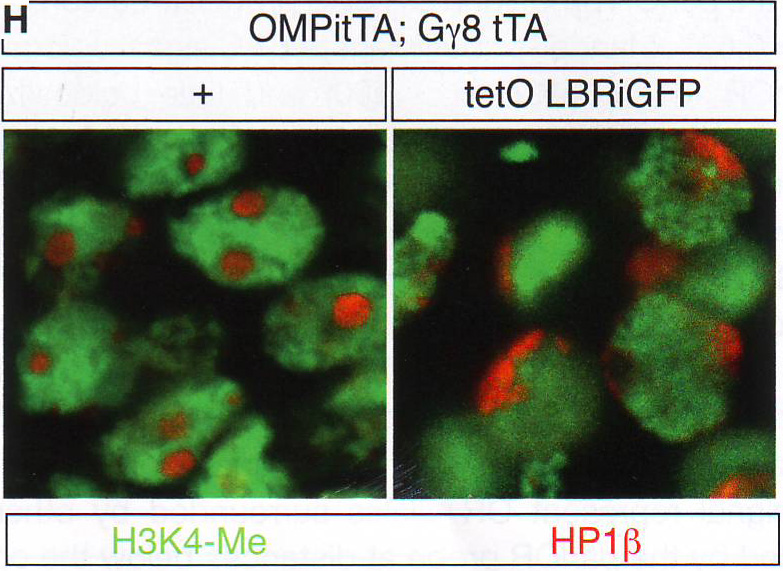
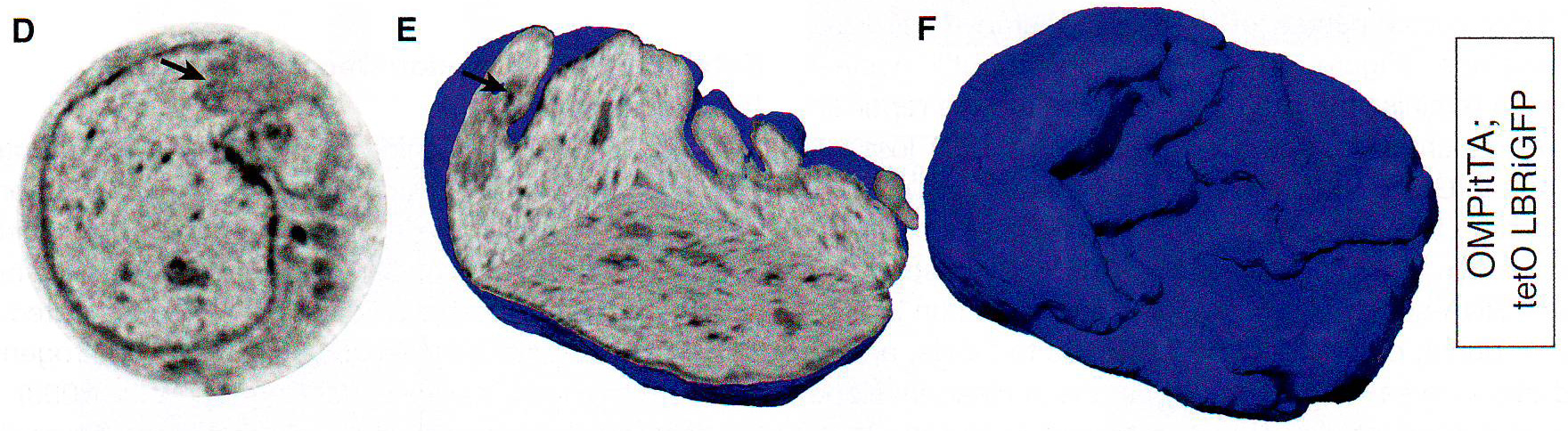
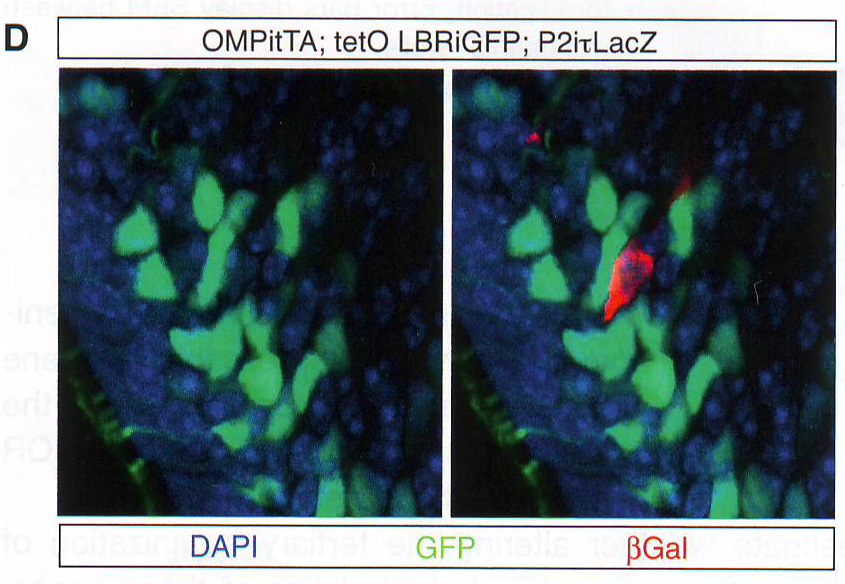
Рис. 1Е-Н. Е. Подсчет количества оптически различимых фокусов, формируемых объединением BAC образцов как на рис 1E в поддерживающих клетках OSN и базальных типах клеток в контроле и ихтиозных MOE отсеках мышей n > 30 для всех групп, p < 0.0001 в группах сравнения между OSN и поддерживающих или базальных клетках в контрольных животных и для сравнения между поддерживающими и базальными клетками в контроле по сравнению с тканями ихтиозных мышей (критерий Стьюдента). F. GFP экспрессия и LBR ИФ в MOE отсеках из tetO LBR - IRES - GFP; OMP - IRES - tTA мыши. Картинка малого увеличения слева показывает GFP сигнал между OE. Изображение высокого разрешения справа показывает GFP (зеленый) и LBR (красный) коэкспрессируются и трансгенный LBR ограничен ядерной оболочкой. G. Изображение ложных цветов DAPI окрашивания в LBR экспрессии OSN по сравнению с контролем OSN показывает потерю OSN-специфичного PH ядра (золотым) при LBR экспрессии. H. ИФ в контрольной и LBR + животных для H3K4-Me1 и HP1 бета показывает реорганизацию OSN ядра при LBR экспрессии.
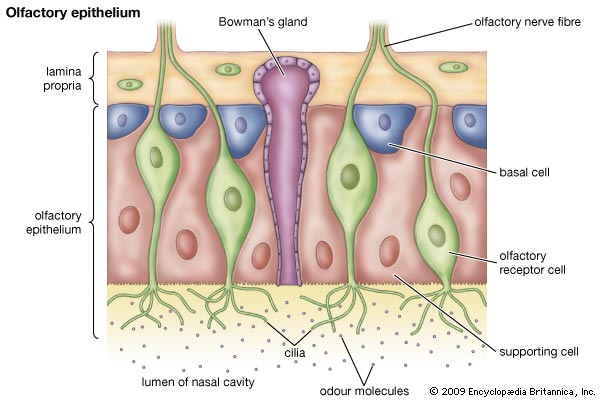
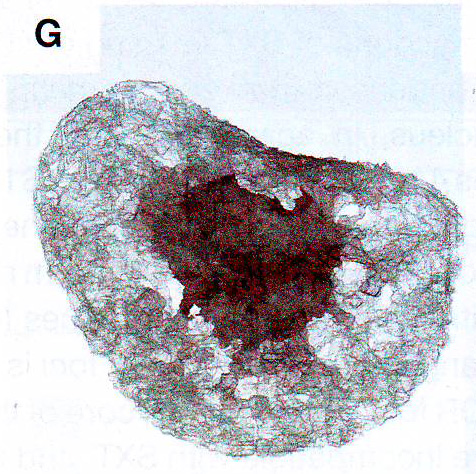
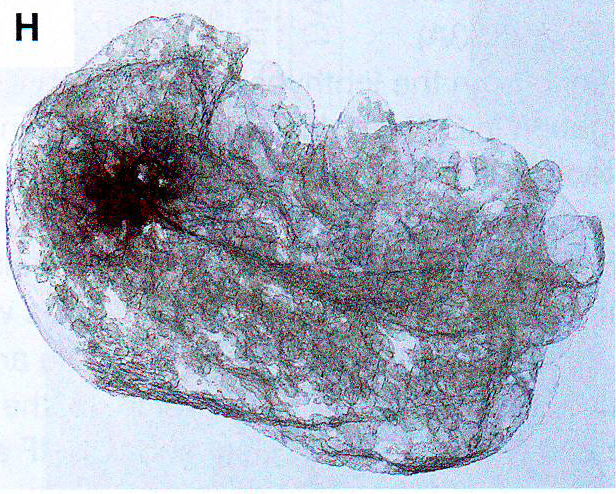
Обонятельный эпителий. Показаны базальные (basal cell), поддерживающие (sustentaculum - supporting cell) и обонятельные нервы Высоко интенсивные точки (более 120 в 8-и битном диапазоне от 0 до 255) были обнаружены только в OSN и отсутствовали в поддерживающих и базальных клетках. Для подсчета распределения панОР сигнала авторы подсчитывали стандартное отклонение интенсивности сигнала во всем пространстве ядра. Среднее стандартное отклонение в OSN 42,3 показало точечное распределение сигнала и 9,3 или 11,3 в базальных и поддерживающих клетках, показало более мягкое распределение (n >100 для каждого типа клеток). ДНК FISH в этом образце в других нервных типах показывает диффузное распределение ОР локусов (данные не показаны, рис S2D), давая дополнительную аргументацию в пользу OSN специфичного ядерного паттерна. Фокальная природа панОР ДНК FISH сигнала предполагает, что ОР аллели из разных ОР кластеров сливаются в строго определенных областях ядра во время OSN дифференцировки. Для проверки этого авторы проверили 10 ОР и 12 неОР BAC пробы и провели двухцветный ДНК FISH c панОР пробой. Наблюдалась экстенсивная колокализация между панОР probe и ОР BAC probes (коэффициент Пирсона 0.637) Mander's коэффициент ВАС сигнала, колокализованного с панОР M1 = 0.835, n > 100) и мало колокализации между панОР probe и не ОР BAC (r = 0.187, M1 = 0.109, n> 100) (рис 1E и 1F и табл S2), предполагая селективность для ОР локусов в составе этих агрегатов (риc S11). Хоть панОР probe включает большую часть ОР локусов, нехватка полного перехлеста между панОР и индивидуальной ОР BAC probe ожидался. ПанОР probe в 200 раз сложнее, чем каждый BAC и вне конкуренции связывается с ОР нацеленной с помощью BAC. Таким образом, BAC сигналы, колокализованные с панОР сигналом представляют ОР аллели, окруженные другими ОР локусами, меченными панОР probe на расстояниях меньших, чем оптическое разрешение конфокальной микроскопии. Комбинированные ОР BAC продуцируют меньше ДНК FISH пятен в OSN (3.94 пятен/ядро/Z stack, n = 38) чем в поддерживающих клетках (9.1 пятен, n = 38) или базальных клетках (8.52 пятен, n = 30), давая независимую верификацию экстенсивной агрегации этих локусов: они оптически видны значительно чаще в OSN чем в базальных или поддерживающих клетках. Не ОР BAC образцы не более агрегированы в OSN (10.08 пятен в OSNs , 6.4 в поддерживающих и 7.1 в базальных клетках, n = 30 для клеток каждого типа). Чтобы исследовать вклад внутри- и межхромосомных взаимодействий в формирование ОР фокусов и колокализацию ОР, авторы использовали два дополнительных пула ОР BAC, один содержащий 7 BAC нацеленных на три кластера на хромосоме 2 и второй содержащий 8 BAC, каждый нацеленный на кластер из разных хромосом. Эти пулы при комбинировании с панОР образцом выявили два уровня организации в OSN: аллели в одном кластере объединяются в оптически однородные сигналы, тогда как кластеры из разных хромосом образуют раздельные сигналы внутри одного и того же панОР фокуса (рис 1G, 1H). BAC из одной хромосомы производят 5,9 точек в поддерживающих клетках и 2,3 точки в OSN (n = 30 для каждой). Тогда как BAC из разных хромосом продуцируют одинаковые количества точек в обоих видах клеток. Однако множественные ОР BAC точки из разных хромосом наблюдались в 50% панОР фокусах и более чем 2 точки на агрегат в 29% клеток (n = 50) (рис 1H). Более того, материнский и отцовский аллели одного ОР кластера остаются в одном ОР агрегате в 6% проверяемых OSN (рис S1H). Итак, панОР фокусы не колокализуются с классами больших повторов перицентромерным хроматином (PH) или другими мультигенными семействами (рис S2A - S2C) и данные не показаны, предполагая, что агрегация ОР кластеров продуцирует четкие и селективные ОР-генные территории.
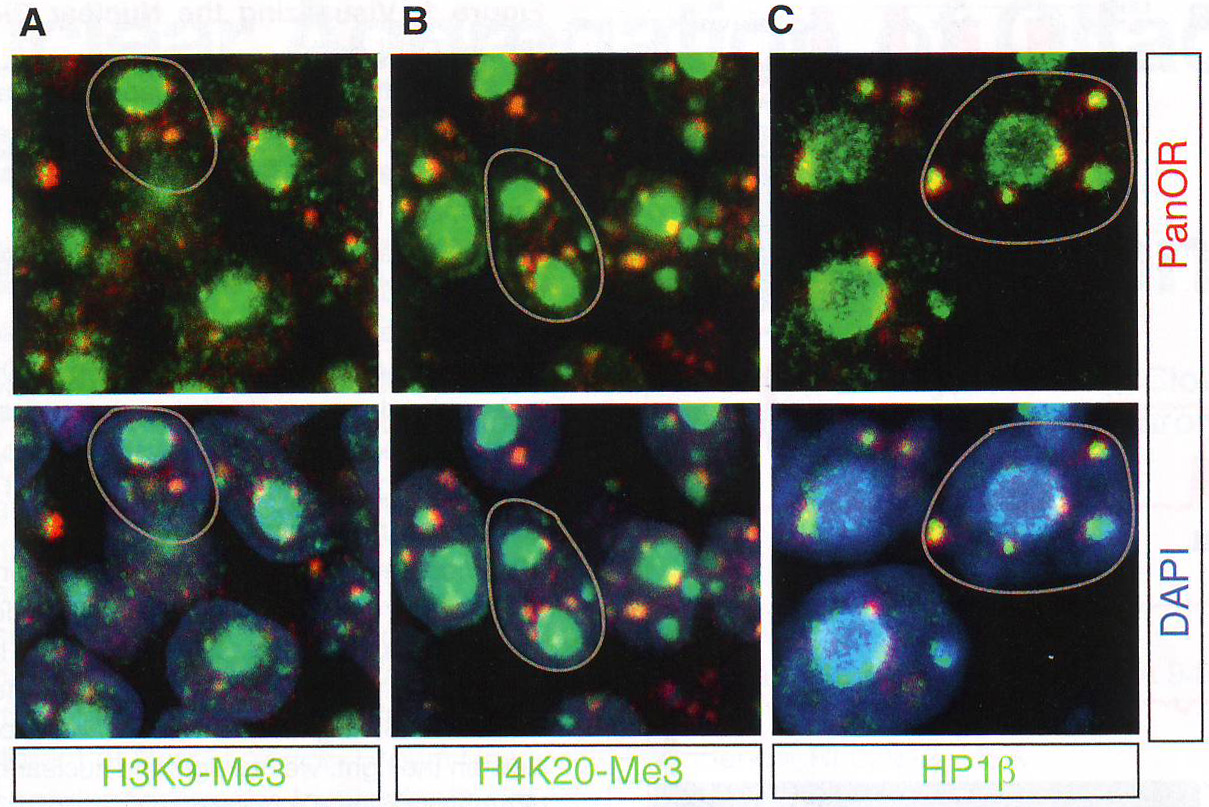
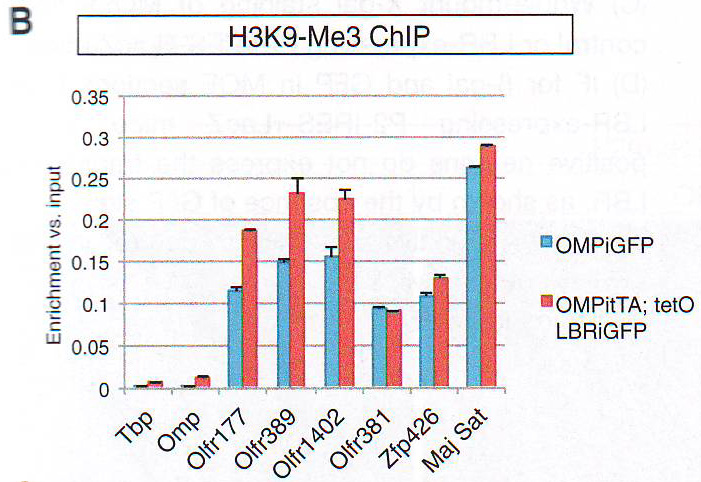
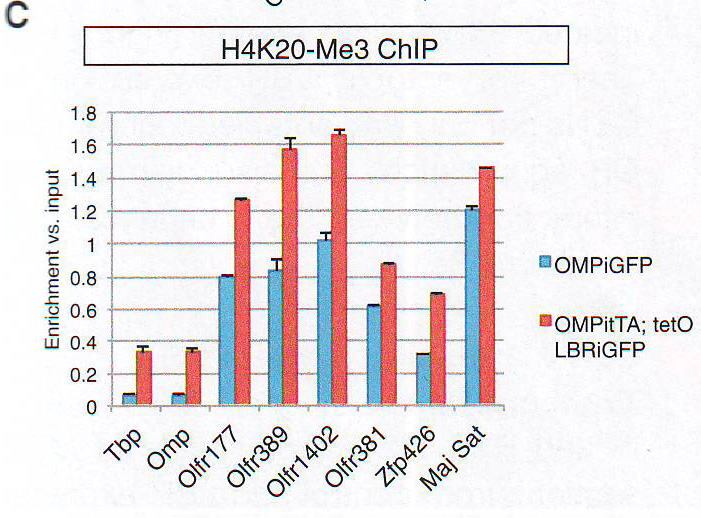
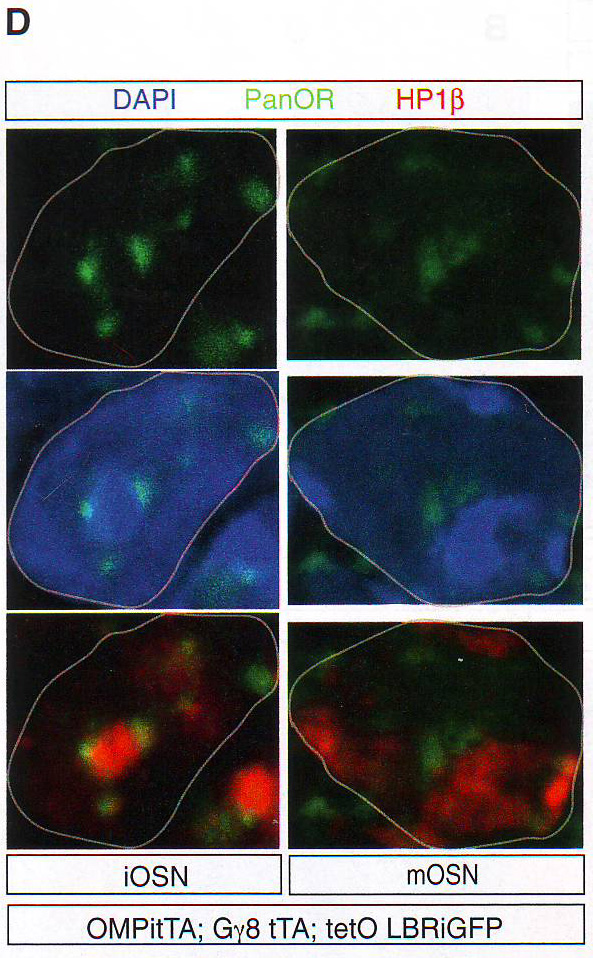
Рис 2А-С. Пространственная сегрегация между активными и молчащими аллелямиДля выявления эпигенетического подтверждения ОР фокусов авторы комбинировали ДНК FISH анализ с иммунофлуоресценцией ИФ против гетерохроматических отметок, находящихся на ОР (Magklara et al., 2011) или гетерохроматин-связывающем белке 1 бета (HP1beta), единственном члене семейства белков HP1, экспрессирующемся в OSN (данные не показаны). Этот анализ выявляет перехлест между ОР фокусами, H3K9-Me3, H4K20-Me3, и HP1beta (рис. 2A - 2C и табл S2), но не с Pol II (рис. S 2E ), состоящей с гетерохроматичной природой этих агрегатов. Эта колокализация дифференцировка-зависимая и специфична для типов клеток, авторы не определили перехлест между двумя сигналами в базальных клетках в MOE или нейронах сетчатки (рис S2D).
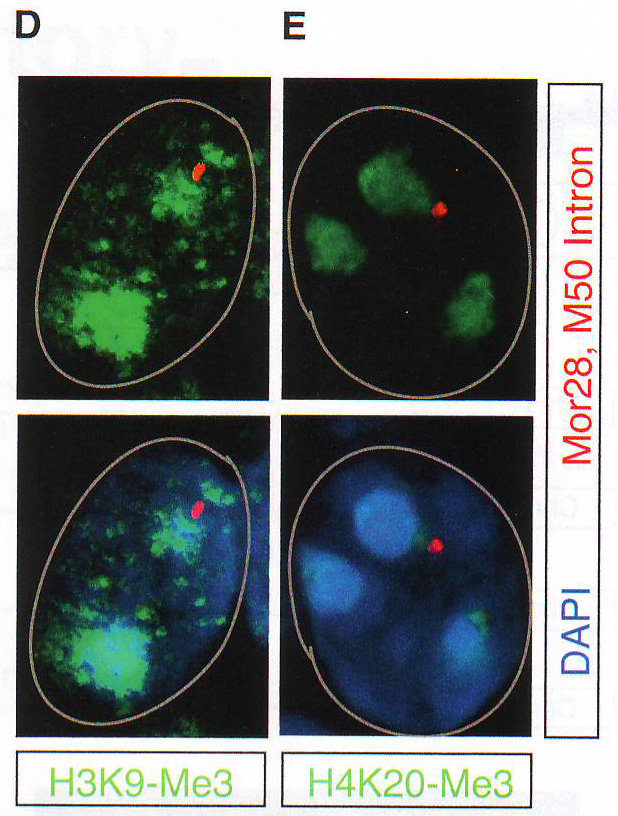
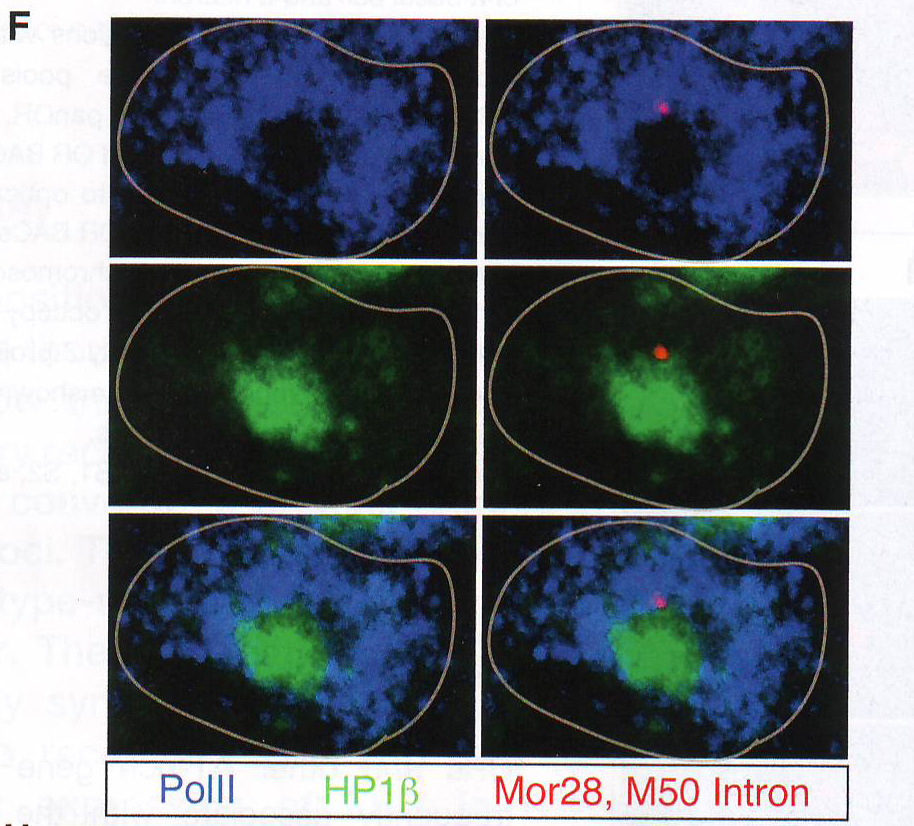
Рис 2D-E. Затем авторы провели образующийся РНК FISH на отсеках MOE с использованием интронных образцов против ОР генов MOR28, M50, M71 и P2 в комбинации с ИФ на НЗК9meЗ, H4K20me3, или HP1 бета. В отличие от интенсивного сигнала панОР активные аллели ОР (H и I) получили 3D поверхностные цветные распределения графиков, отвечающих за клетки от F и G соответственно. Свечение изображения интерпретировали как высоту графика. Образующиеся ОР транскрипты показаны красным. HP 1 бета фиолетовым и Pol II (H) или H3K27 ацетил (I) зеленым.
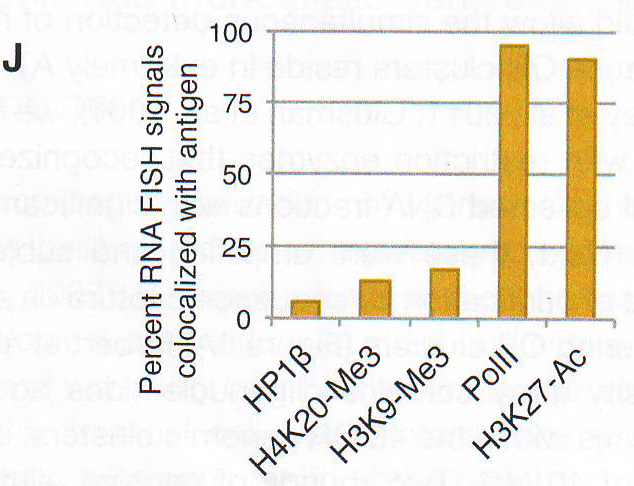
Рис 2F-J. Ручной подсчет колокализации образующихся транскриптов и антигенов представлен на D - G. Сигналы подсчитывали как колокализованные, когда было некоторое перекрывание между образовавшимися транскриптами и антигеном. n = 150 для HP 1 бета , 31 для H4K20me3, 12 для H3K9me3, 64 для Pol II , и 64 для H3K27-AC .Наблюдался малый перехлест с любой из трех гетерохроматиновых меток (рис 2D -2J, S2F, и S2G и табл. S2). Авторы также комбинировали образующиеся РНК FISH с ИФ для Pol II или H3K27-ацетил и H4K20me3 или HP 1 бета (рис 2F -2J и табл S2). Этот эксперимент подтверждает, что активные ОР аллели пространственно сегрегированны от молчащих ОР аллелей и находятся в области эухроматина. LBR (рецептор ламина b) организует топологию ядер OSN (обонятельных чувствительных нейронов)Интриует то, что большинство ОР генов и PH находятся рядом с центром OSN ядер вместо того, чтобы быть распространенными в области ядерной оболочки. (рис S1A) . Эта морфология ядра «наизнанку» напоминает архитектуру ядра, описанную в гомозиготных ихтиозных мышах, спонтанных LBR мутантах с потерей функции гена (Goldowitz and Mullen, 1982). LBR – белок ядерной оболочки, который взаимодействует с HP 1 бета и гетерохроматином (Hoffmann et al ., 2002; Okada et al ., 2005; Pyrpasopoulou et al ., 1996). РНК последовательность выявила постоянное уменьшение уровня мРНК гена LBR во время дифференцировки от HBC до OSN и ИФ подтверждено, что тогда как LBR присутствует в ядерной оболочке базальных и поддерживающих клеток, он отсутствует в нейрональной культуре клеток основного обонятельного эпителия (МОЕ) (рис ЗА , 3B, и S3A).
Рис 3А.
Рис 3B. панОР ДНК FISH на MOE срезах ихтиозных мышей не выявило изменений в ОР агрегации в OSN , которой уже и так не хватает в LBR . Однако архитектура ядра и ОР организация ихтиозных базальных и поддерживающих клеток приближается к дикому типу OSN (рис. 3C , 3D , и S3B).
Рис 3C.
Рис. 3D. PH образует большие расположенные в центре фокусы в обоих типах клеток, а ОР образует агрегаты на периферии перицентромерной области. Согласно объединенному BAC исследованию на ихтиозных мышах количество ДНК FISH пятен однородно в трех типах клеток и базальные и сустентакульные клетки имеют схожее количество ДНК FISH пятен для контроля OSN (рис 3E), поддерживая роль LBR снижения регуляции в агрегации ОР.
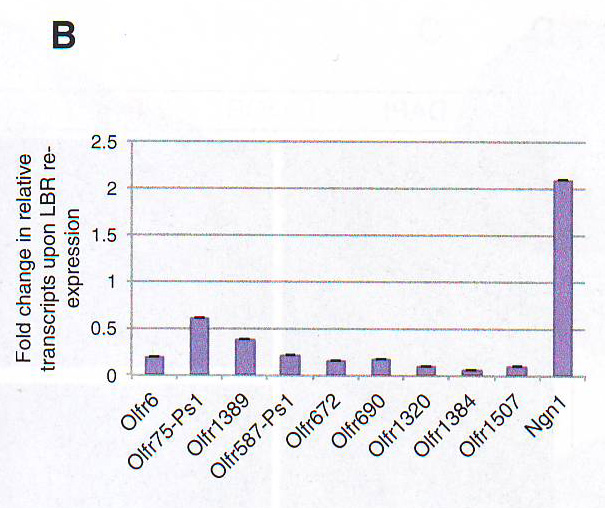
Рис 3Е. Так как эктопическая агрегация происходит в двух типах клеток, которые не экспрессируют ОР, и вероятно не содержат факторы транскрипции, отвечающие за активацию ОР, эффект этой мутации на ОР экспрессию и OSN таргетинг маловероятен и не был обнаружен (данные не показаны, рис S3C). Поэтому авторы решили провести противоположный эксперимент: для восстановаления LBR экспрессии в OSN вместо удаления LBR из клеток, которые не экспрессируют ОР. Авторы вывели tetO LBR - IRES - GFP трансгенную мышь, которую они скрестили с OMP - IRES - tTA мышами, чтобы добиться экспрессии LBR в OSN. Одна трансгенная линия экспрессирует трансген в значительной пропорции с OSN (рис 3F ). Как и эндогенный LBR, трансгенный LBR ограничен ядерной оболочкой без диффундирования в нуклеоплазму (рис 3F). Авторы использовали эту трансгенную линию для анализа эффекта эктопичной экспрессии LBR на морфологию ядра OSN.
Рис. 3F DAPI окрашивание становится менее интенсивным и PH перемещается к периферии ядра LBR и OSN (рис 3G и S3D). Обонятельные нейроны в этих срезах, которые не экспрессируют трансген, имеют морфологию ядра близкую к дикому типу (рис S3D). ИФ показывает HP1бета вовлечение в ядерную оболочку в LBR + OSNs, тогда как центрально сдвинутый эухроматин занимает большую часть ядра. (рис. 3H, S3E, и S3F). Таким образом эктопичной LBR экспрессии в постмитотичной клетке достаточно для того, чтобы перевернуть «наизнанку» установку и вовлечь PH в ядерную периферию.
Рис 3G.
Рис 3Н. Эктопичная экспрессия LBR деконденсирует гетерохроматин в OSNИФ не дает информации о структурных и биофизических изменениях, происходящих в OSN хроматине при эктопической экспрессии LBR. Чтобы получить эту информацию авторы зафиксировали контрольные и LBR + OSN мягкой рентгенографией (SXT), метод высокого разрешения применимый к полностью гидратированным, нефиксированным и не окрашенным клеткам и измеряет концентрацию углерода и азота в биологическом образце (McDermott et al ., 2009). Orthoslices (компьютер-генерированные срезы) и 3Д реконструкции SXT изображений контрольных OSN выявили, что более конденсированный (темный) хроматин расположен в центре ядра, что согласуется с морфологией, наблюдаемой при ИФ (рис 4A, 4B, и 4G и видео S1). Авторы также обнаружили чрезвычайно плотные структуры по периферии PH ядра, что специфично для этого типа клеток. Только ядра сперматозоидов имеют частицы хроматина с более высоким уровнем упаковки (данные не приведены). Хотя расположение этих темных фокусов сходно с расположением ОР фокусов вокруг PH центра OSN ядра ДНК FISH или ИФ несовместимы с SXT и поэтому невозможно прямо доказать, что это идентичные структуры.
Рис 4А - С. OMPiGFP. SXT изображения LBR + OSNs показывает перераспределение наиболее конденсированного хроматина к ядерной мембране (рис 4D, 4E, и 4H и видео S2).
Рис 4D-F.
Рис 4G
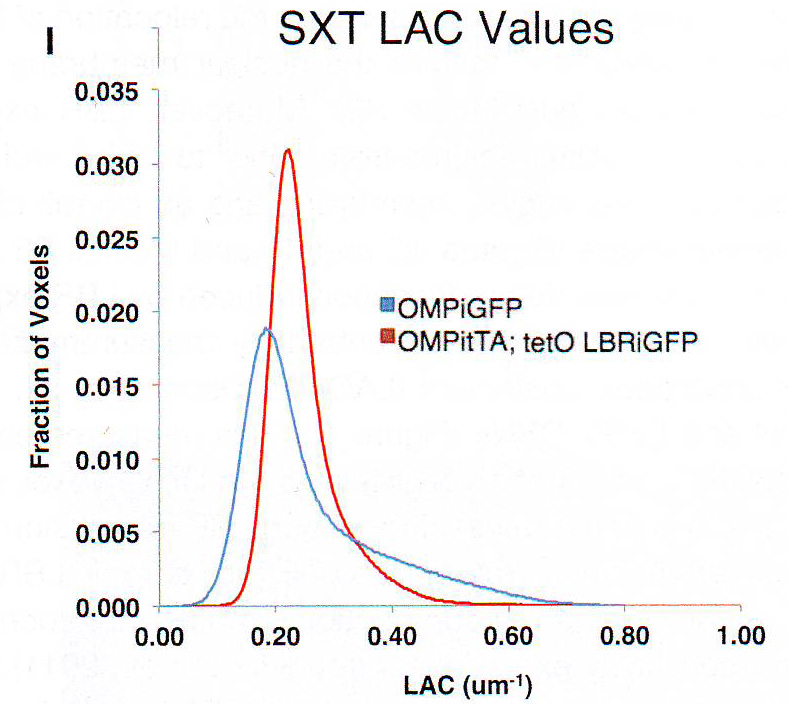
Рис 4H. Кроме того, экспрессия LBR увеличивает объем ядра с 103 до 135 объемных единиц (u) и вызывает сворачивание ядерной мембраны и всеобщее изменение формы ядра (рис 4C и 4F и видео S3 и S4). В итоге деконденсация хроматина, вызванная экспрессией LBR в OSN количественно описана изменением коэффициента линейной абсорбции (LAC). ( McDermott et al ., 2009) контроля и LBR + OSNs (рис 4I)
Рис 4I. Это измерение, которое отображает концентрацию органического материала на voxel (воксел - минимальный адресуемый объёмный элемент изображения трёхмерного пространства), подтверждает потерю плотности самыми плотными структурами при экспрессии LBR. Таким образом, если конденсированные области соответствуют ОР фокусам, эктопичная экспрессия LBR должна вызывать декомпактизацию ОР гетерохроматина. ДНКазные эксперименты чувствительности (Magklara et al., 2011) в ядре от сортированных с помощью флуоресценции активированных клеток (FAC-сортированные) контроль или LBR + OSNs подтверждает значимость декомпактизации ОР и перицентромерного гетерохроматина при LBR экспрессии (Рис 4J).
Рис 4J. Эктопическая экспрессия LBR нарушает агрегацию обонятельных рецепторовПространственная реорганизация HP1 бета, удаление темных фокусов, обнаруживаемых путем SXT и повышение ДНКазной чувствительности ОР хроматина предполагают, что эктопическая экспрессия LBR нарушает агрегацию локусов ОР. Чтобы проверить это, авторы провели FISH анализ ДНК с панОР образцом в секции LBR экспрессирующих трансгенных мышей. Изображения низкого разрешения показывают значительные эффекты эктопической экспрессии LBR на распределение локусов ОР. В верхушечном LBR + нейрональном слое интенсивные ОР фокусы растворяются, в отличие от незрелых OSN и предшественников, которые пока еще не экспрессируют трансген, но уже подавили эндогенный LBR остаются фокально ОР структурированы (рис 5А и S4A ).
Рис 5А. Чтобы изучить, как повреждение третичной организации ОР локусов влияет на эпигенетическую характеристику этих генов, авторы исследовали ассоциацию ОР генов с H3K9me3, H4K20me3, и НР1 бета в LBR + OSN . НЗК9meЗ и H4K20me3 оставались обогащенными ОР локусами при экспрессии LBR в нативных ChlP - qPCR эссе на FAC -отсортированных OSN. (Рис 5B и 5C ) и FISH -ИФ (рис S4B и S4C и Табл S2).
Рис 5B.
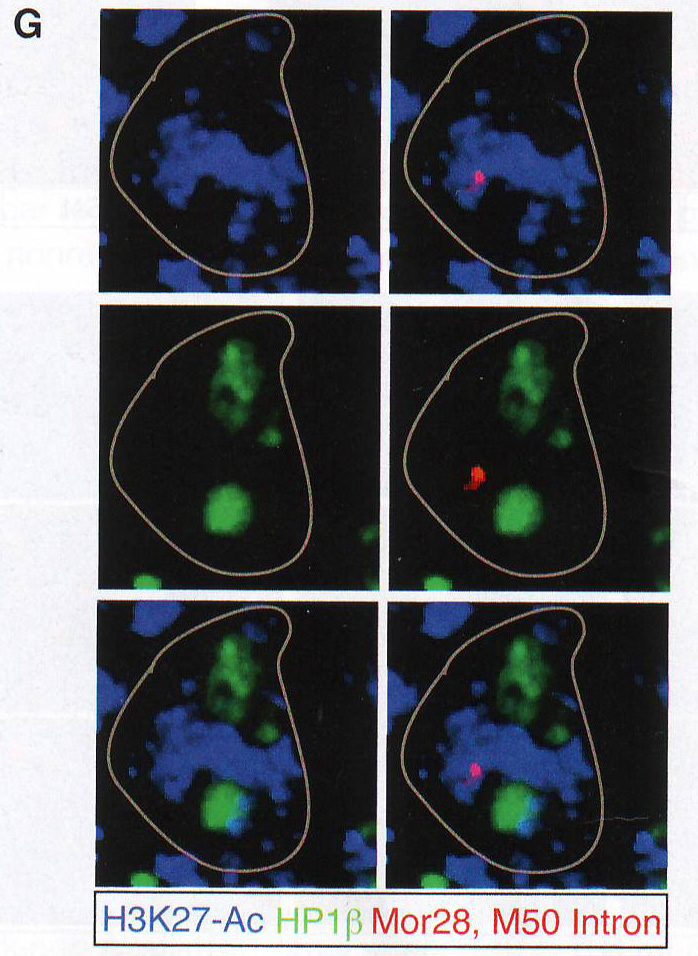
Рис 5C. И наоборот, ассоциация ОР локусов с HP1 бета была снижена при измерении с помощью FISH ИФ (Рис 5D и Табл S2).
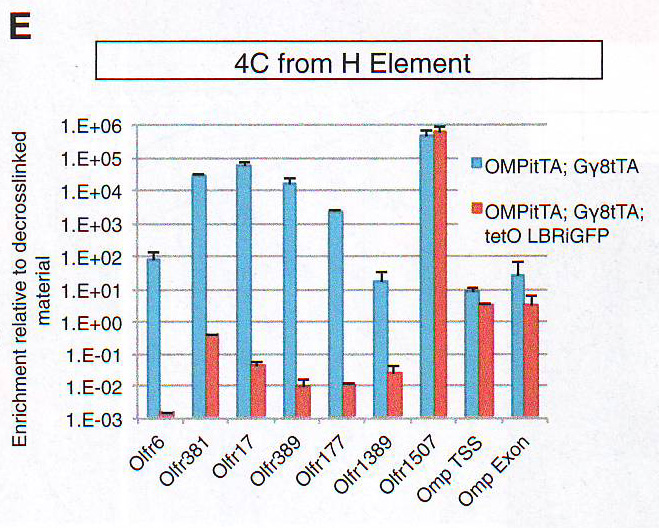
Рис 5D. Снижение перекрытия между H4K20me3 и HP1 бета также наблюдалось в LBR + OSN. Таким образом, несмотря на сохранность гетерохроматиновых гистонных меток ОР теряют свою агрегационную структуру (порядок) и свою негистоновую гетерохроматиновую оболочку при LBR экспрессии, которая сохраняется при повышенной ДНКазной чувствительности. В диком типе OSN активные ОР аллели взаимодействуют с H энхансером. Для проверки действительно ли экспрессия LBR аннулирует межхромосомные взаимодействия между активным аллелем и Н, авторы провели фиксацию конформации циркулярной хромосомы (circularized chromosome conformation capture) (4С), используя обратный Н ПЦР праймер, как описано в работе Lomvardas et al ., 2006 на LBR экспрессирующих или контрольных MOE. Для увеличения количества экспрессирующих LBR клеток в этой микс-популяции авторы комбинировали два tTA драйвера (OMP - IRES - tTA и Gy8 tTA). Обогащение различных ОР сиквенсов в этой 4С библиотеке было исследовано путем количественного ПЦР. Экспрессия LBR в OSN приводит к потере большинства связей H-ОР. В LBR трансгенах Н остается связанным только со связанным ОР MOR 28 (Olfr 1507), расположенным 75Кб вперед по цепи (рис 5Е). Поэтому эктопичная экспрессия LBR в OSN не только предотвращает агрегацию гетерохроматина, но и нарушает взаимодействие между Н энхансером и не связанными ОР-ами.
Рис 5Е.
Экспрессия LBR ингибирует транскрипцию обонятельных рецепторовИФ и РНК FISH эксперименты в MOE срезах клеток контрольных и LBR экспрессирующих мышей выявили трехкратное снижение количества нейронов, экспрессирующих определенные ОР у трансгенных мышей (рис 6А).
Рис 6А.
Рис 6B.
Рис 6С. Важно, что большинство нейронов, в которых остается высокий уровень экспрессии ОР, не экспрессируют LBR (данные не приведены, рис 6D).
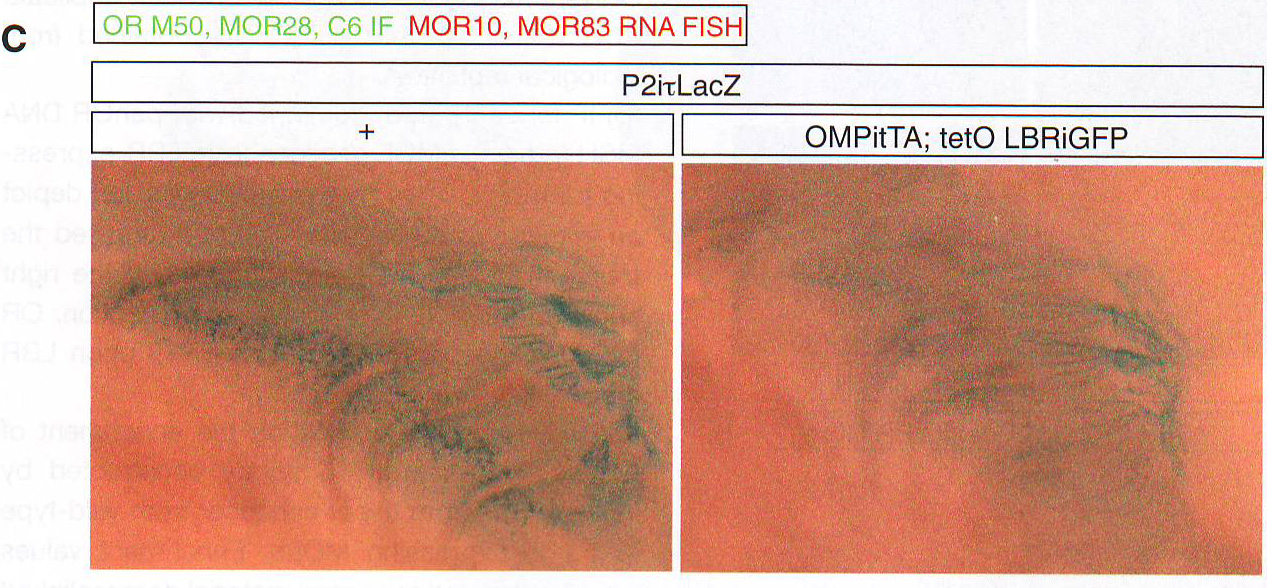
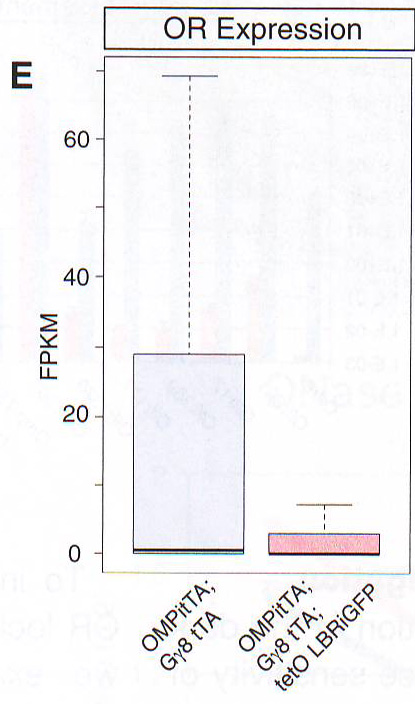
Рис 6D. Для количественного измерения эффекта LBR на экспрессию ОР авторы использовали FAC сортировку для изоляции контрольных и LBR+ OSN и провели количественный ПЦР обратной транскриптазой (qRT - PCR). Этот анализ дал те же результаты значительного подавляющего эффекта на экспрессию ОР (рис 6В). Схожим образом полно количественное X - gal (BCIG - 5-bromo-4-chloro-indolyl-?-D-galactopyranoside) окрашивание в МОЕ P2- IRES - TLacZ мышах, скрещенных с LBR -экспрессирующими трансгенными мышами показывает снижение X-gal сигнала, поддерживая гипотезу о том, что LBR экспрессия имеет ингибирующий эффект на ОР экспрессию (рис 6С). Нейроны, в которых сохранялся высокий уровень транскрипции p - gal белка обычно не экспрессировали LBR трансген, как показано на ИФ при p - gal и GFP на срезах этих мышей (рис 6D). Так как OMP запускает экспрессию LBR только после выбора ОР, этот результат показывает пост-выборную снижающую регуляцию этого 2Р аллеля и оставшийся ОР репертуар. Чтобы проверить, применим ли ингибиторный эффект LBR к генам, которые не следуют пространственной регуляции ОР генов, авторы использовали трансгенные ОР, которые находились под контролем тетО промотера ( tetO MOR 28- IRES -- rLacZ ). Этот трансген также нес НЗК9meЗ и H4K20me3 (данные не приведены), но в отличие от эндогенных ОР их гетерохроматинизация не OSN специфична и возможно вызвана их многочисленными вставленными копиями (16 тандемных копий) (Garrick et al ., 1998). Этот трансген не взаимодействует с эндогенными ОР и Н энхансером (рис H11 и данные не приведены). В согласии с репрессирующей значимостью этого трансгена его экспрессия спорадическая при скрещивании с OMP - IRES - tTA драйвером, но частота повышается при скрещивании с LBR экспрессирующими трансгенами (рис S5B). Это соотносится с простым способом генной регуляции, при которой декомпактизация хроматина позволяет tTA связаться с tetO промотером в большем количестве клеток и активация транскрипции происходит чаще, так как прямолинейная модель не применима к эндогенным ОР. Чтобы определить эффект LBR экспрессии в масштабах всего генома в DSN авторы провели РНК-сиквенс из всех МОЕ препаратов. Чтобы увеличить содержание LBR + OSN в этой смешанной популяции авторы комбинировали два tTA драйвера OMP - IRES - tTA и Gy8 tTA . В соответствии с наблюдениями в FAC сортирующими нейронами LBR экспрессия в OSN вызывает восьмикратное снижение полной ОР экспрессии (рис 6Е).
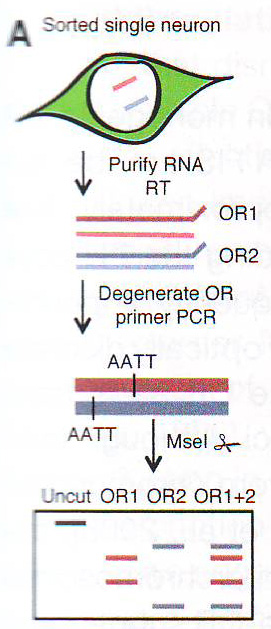
Рис 6Е. Хотя ОР экспрессия снижена, большинство генов, определяемых в OSN (около 14000 генов) не подвержены влиянию эктопической экспрессии LBR (367 не- OR транскриптов значительно подавляются и 873 транскрипта значительно активизируются, Cuffdiff FDR 0.05, анализ генов проведен по крайней мере по 10 ридов). Интересно то, что экспрессия некоторых маркеров незрелых OSN и предшественников клеточных популяций повышена у этих животных (рис S5A). В общем масштабе эти изменения предполагают частичное изменение в сторону меньшей дифференцировки, нежели чем удаление зрелых OSN, как показано путем недостатка повышения апоптоза в трансгенных МОЕ (рис S5C). Эктопическая экспрессия LBR нарушает сингулярность транскрипции обонятельных рецепторовПонижающая регуляция транскрипции ОР в LBR+ нейронах интуитивно подразумевает, что доступность хроматина ОР увеличивается и что эти локусы сняты с HP 1 бета. Разрушение долгосрочных взаимодействий с активирующими энхансерами, например, Н, может способствовать этой понижающей регуляции. Более того, хотя общая декомпактизация ОР хроматина может сделать все ОР аллели транскрипционно компетентными, возможно, что OSN не могут поддерживать транскрипцию более 2800 аллелей ОР на уровне одиночно транскрибируемых ОР. Чтобы проверить это, авторы провели ПЦР обратной транскриптазой одной клетки с дегенерированными ОР праймерами, затем провели рестрикцию ферментом и электрофорез (Buck and Axel , 1991; Рис 7A и S6A ).
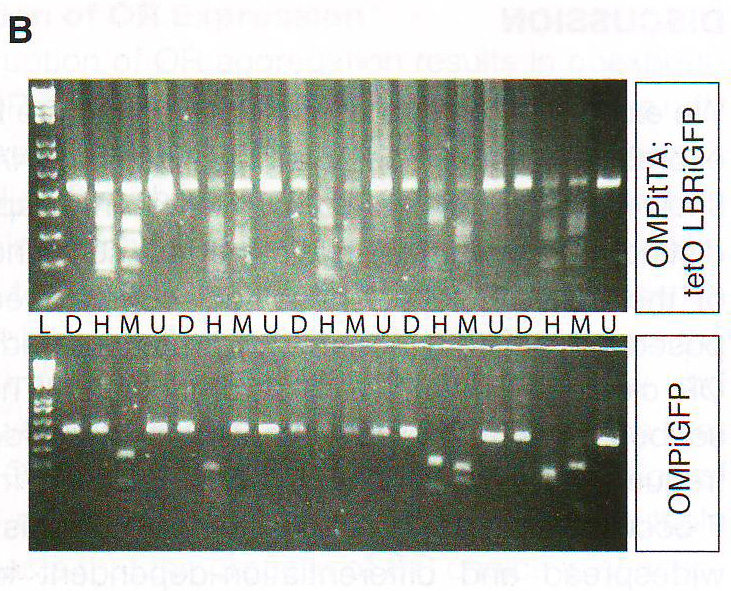
Рис 7А. Авторы получили 10 кДНК библиотек с одной клетки с каждого генотипа (см. описание проведения эксперимента) и проверили представленность ОР Dral, Hinfl, и Msel расщеплением с помощью соответствующих ферментов рестрикции. Строение дегенерированных ОР ампликонов отличается у OSN и LBR+ OSN . В каждом OSN LBR+ сумма пар оснований одиночных продуктов расщепления превосходит длину нерасщепленного ПЦР продукта, тогда как контрольные ампликоны содержат только 1 продукт. (рис 7B , показаны 5 ампликонов каждого генотипа. Схожие результаты были получены для пяти остальных ампликонов и расщеплением Mbol – данные не приведены).
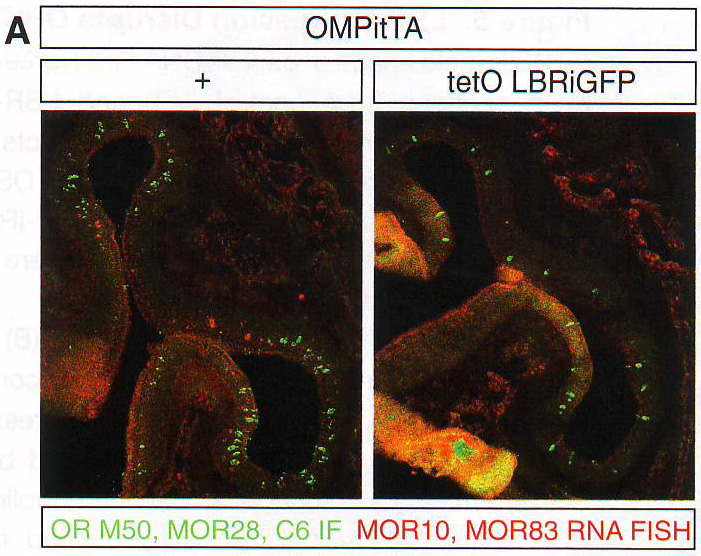
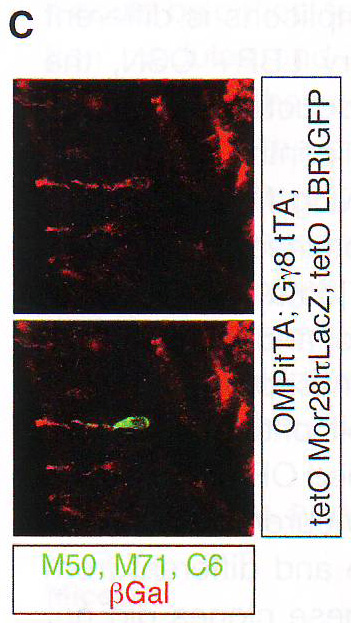
Рис 7B. Секвенирование 10 клонов каждого из двух контролей, двух LBR экспрессирующих ампликонов подтверждает, что ОР транскриптом более сложно устроен в LBR+ OSN. В обоих контрольных ампликонах все 10 клонов были идентичны и сиквенс клонированного ОР соответствовала паттерну расщепления. В случае LBR+ нейронов 10/10 и 9/10 клонов каждого ампликона были уникальны и отличались друг от друга. Более того, последовательности этих клонов не соответствовали паттерну расщепления, из чего можно предположить, что эти библиотеки очень сложно устроены и наблюдаемые четкие группы представляют мигрирующие вместе группы сходного размера, расщепленные из большого количества различных ОР. Чтобы проверить это, авторы секвенировали 96 независимых колоний из третьей LBR+ одноклеточной библиотеки и обнаружили 46 различных ОР из 11 хромосом без всякой зональной рестрикции (табл S3). Таким образом, экспрессия LBR в OSN нарушает правило «один рецептор на один нейрон» и вызывает коэкспрессию большого количества ОР. Чтобы опровергнуть предположение о том, что множественные ОР экспрессируются в малых количествах также в контрольных OSN, но скрыты высоким уровнем экспрессии выбранного аллеля, авторы провели обратно-транскриптазную реакцию одинаковых объемов, полученных из одной клетки из контрольного и LBR+ нейрона и провели дегенеративный ПЦР и расщепление. Хотя ОР ампликон из контрольного нейрона доминирует в реакции, эктопически коэкспрессируемые ОР из LBR экспрессирующего нейрона все еще были определяемы после расщепления (рис S6B). В дополнение к ПЦР подтверждению коэкспрессии ОР в LBR+ нейронах авторы провели ИФ для ОР М50, М71 и С6 в tetO - LBR / tetO - MOR28 двойных трансгенах. Большинство OSN, в которых оставался определяемый уровень ОР на ИФ были LBR негативны, поэтому в этих OSN коэкспрессия ОР крайне редка. Чтобы преодолеть это, авторы использовали частую экспрессию трансгенных MOR28 в LBR экспрессирующих мышах. ИФ в МОЕ срезах у двойных трансгенов выявила двойные положительные OSN (рис 7С), которые не обнаруживаются в отсутствии LBR , как показано ранее ( Fleischmann et al ., 2008; Nguyen et al ., 2007). Наиболее вероятно, эти OSN закрывают эндогенные ОР в ответ на экспрессию LBR, когда активируют деконденсирующиеся трансгенные ОР.
Рис 7С. Особенности экспрессируемых ОР аллелей дают инструкции целевым нейронам на один клубочек ( Mombaerts , 2006). По этой причине авторы исследовали прицельно экспрессию аллеля P2- IRES - i - LacZ в нейронах LBR трансгенов путем ИФ для p - gal в срезах обонятельной луковицы, p - gal белок стабилен в аксонах еще долго после угасания цитоплазматического сигнала (рис S6D); поэтому можно использовать этот подход к исследованию целевого влияния LBR экспрессии. В группе контрольных мышей p - gal положительные волокна объединяются в четкие клубочки (рис 7D), где очень мало аксонов направлены в неправильные гломерулы. (крайний пример приведен на рисунке S6C ).
Рис 7D Однако при экспрессии LBR p - gal положительные волокна распространяются к необыкновенно большому числу клубочков (около 30 на полушарие у одной крысы). Авторы определили экстра четкие клубочки и блуждающие волокна возле клубочков 2P дикого типа и в эктопической позиции в луковице (рис 7D и S6C). ОбсуждениеАвторы исследовали роль архитектуры ядра при моногенной экспрессии обонятельных рецепторов. Используя комплексный ОР-специфичный ДНК FISH образец, они показали, что ОР гены сливаются в примерно пять четких и кажущихся эксклюзивными фокусов, окружающих периферический хроматин ядер OSN. Эти фокусы содержат часто наложенные один на другой локусы ОР с одной и той же хромосомы и оптически неразличимые ОР кластеры разных хромосом. ОР аллель, транскрибируемая в каждом OSN отсутствует в этих фокусах. Хотя низкочастотные взаимодействия между кластерами ОР на 7-й хромосоме случаются в печени и мозге эмбриона (Simonis et al ., 2006), широко распространенная и дифференцировка-зависимая межхромосомная агрегация и фокальная организация всего ОР субгенома может быть уникальна для OSN клеток-потомков. Поэтому наши эксперименты предполагают, что первичная эпигенетическая характеристика подкреплена вторичной и третичной репрессивной организацией: внутрихромосомное уплотнение (компактизация) и межхромосомная агрегация ОР генов в OSN. Важность в этой детальной системе показана на примере разрушения ее (этих агрегатов), что приводит к повреждению моногенной ОР транскрипции и коэкспрессии большого количества ОР. LBR и периферический хроматин как организаторы агрегации обонятельных рецепторовМутация с потерей функции LBR приводит к эктопичной агрегации ОР в базальных и поддерживающих клетках. Противоположно этому экспрессия LBR в OSN изменяет морфологию ядра и разрушает фокусы ОР. Поэтому регуляция ОР экспрессии управляет пространственной агрегацией генов ОР в МОЕ. LBR мог бы действовать напрямую на ОР (связываясь с HP 1 бета) и косвенно, вовлекая перицентромерный гетерохроматин в ядерную оболочку. ОР не вовлекались в ядерную оболочку так же эффективно, как PH. Меньший размер ОР кластеров и их геномное расположение в эухроматине могло бы сделать их менее подвижными, чем акроцентричный PH, который вовлечен в основном в периферическую ядерную оболочку. Кроме того, перемещение гена к ядерной оболочке требует разделения клетки (Zullo et al., 2012) что не происходит в клетках OSN. В любом случае, в диком типе OSN PH может предоставить платформу, на которой образуются ОР агрегаты после снижающей регуляции LBR и в LBR+ OSN нейронах релокация PH может помочь в разрешении ситуации построения ОР агрегатов. Конечный биохимический выход этих перестановок – это декомпактизация ОР гетерохроматина, демонстрирующая снижение LAC величины в SXT и повышение ДНКазной чувствительности. Неспецифичный эффект эктопической экспрессии LBR не может быть исключен, хотя большая часть не ОР генов не задета этой эктопической экспрессией. Известные и предполагаемые гены, активирующие ОР экспрессию, например, члены семейства Emx 2, Lhx 2 и Ebf (Fuss and Ray, 2009), также повышают регуляцию или не затронуты экспрессией LBR. (рис. S5A), делая вторичные эффекты маловероятной причиной снижающей регуляции ОР. Кроме того, слабая ферментная активность LBR, который производит эргостерол, не должен участвовать в регуляции ОР, так как ихтиозные мыши не имеют дефицита ОР экспрессии. Кроме того, ферменты, которые продуцируют субстрат для LBR оба экспрессируются на очень низком уровне в OSN (данные не приведены), поэтому LBR не является ферментом, определяющим уровень активности этого метаболического пути, и его повышающая регуляция не повлияет на уровень эргостерола. Пространственная регуляция экспрессии обонятельных рецепторовФакт, что разрушение агрегации ОР приводит к коэкспрессии многих ОР генов показывает, что эта организация критична для эффективного глушения не выбранных ОР аллелей в каждом OSN. Также предполагается, что гетерохроматиновые метки, найденные на ОР, которые остаются обогащенными в этих локусах, при индукции LBR не достаточны для предотвращения базовой транскрипции в отсутствии упаковки более высокого порядка этих областей хроматина. Как при феномене транскрипционного задавливания (Gill and Ptashne, 1988) авторы сделали интуитивное наблюдение, что индукция LBR в OSN вызывает значительное общее снижение транскрипции ОР, позволяя одновременную коэкспрессию множества аллелей. Это наводит на мысль, что процесс выбора ОР концептуально более сложен, чем, например, регуляция tetO MOR28 трансгена и большое количество аллелей ОР может быть значительным фактором. Авторы предполагают, что беспрецедентное количество генов, разделяющих одинаковые факторы транскрипции (Clowney et al ., 2011) образуют эффективное скрытие большей части этих аллелей, в то время, как один из аллелей транскрибируется значительно больше всех остальных. Таким образом, гетерохроматинизация большинства ОР локусов и их агрегации в большие ядерные фокусы не только обеспечивает их эффективное глушение, но также скрывает тысячи мест связывания факторов транскрипции, которые отделяют активирующие белки от выбранных аллелей. В итоге идентификация множества ОР в каждом LBR+ нейроне может отражать последовательный процесс переключения (Shykind et al ., 2004), вызванный понижающей регуляцией изначально выбранного ОР и неспособности сделать новый продуктивный выбор ОР. Геномная конкуренция может быть не единственной причиной для понижающей регуляции ОР при LBR экспрессии. Одинаково тщательно продуманная схема межхромосомных взаимодействий может быть вовлечена в активацию одного ОР аллеля. Следовательно уход от гетерохроматических фокусов может быть недостаточным для активации транскрипции ОР. ОР ген может нуждаться в перемещении в специальный транскрипционно-компетентный центр меж хромосомами, как в случае с активацией IFN бета (Apostolou и Thanos, 2008). С этим также согласуется факт, что активный ОР аллель часто находится прикрепленным к гетерохроматическим фокусам. Это может значить, что агрегация ОР не только приглушает ОР аллели, но также организует некоторые из них – возможно, находящиеся на периферии фокусов – для активации. Баланс и организация агрегации ОР для будущей активации может объяснять причину селективности этих фокусов для последовательности ОР. Таким образом, тщательный пересмотр ядра, вызываемый экспрессией LBR может также разрушить взаимодействия между энхансерами длинного расстояния и выбранного ОР аллеля, что приводит к снижению регуляции ОР. Наблюдение, что LBR экспрессия нарушает транс взаимодействие между Н и ОР соотносится с данной моделью. Хотя нет генетических доказательств, где требуются простые транс взаимодействия для ОР транскрипции (Khan et al ., 2011) более тщательная схема межхромосомных взаимодействий может управлять ОР активацией, как в случае с приглушением ОР. Ядерная реорганизация во время развитияРазличия в ядерной топологии могут наблюдаться во многих сенсорных эпителиях (данные не приведены) и регуляция экспрессии LBR может управлять некоторыми из них. Хотя реорганизация ядра может служить дополнительным функциям (Solovei et al ., 2009) она может быть критична для выполнения тканеспецифичной дифференцировки и может постоянно выключать программы генной экспрессии, когда они проявляются. Таким образом, на самом высоком уровне организации хроматина эпигенетический «ландшафт» становится физическим ландшафтом, где определенные гены и регуляторные последовательности спрятаны или выставлены в согласии с типом клетки и ее функцией. Будущие эксперименты определят, сходны ли механизмы пространственной регуляции описанным в этой статье применимы к менее решающим направлениям развития клетки, не вовлекая процесс выбора одного из тысячи аллелей. Описание экспериментальных процедурМыши Мышей содержали в стандартных условиях в согласии с правилами IACUC как описано ранее (Magklara et ai ., 2011). РНК FISH эксперименты были проведены на 6 – 10 день после рождения (p6 – p10). ДНК FISH эксперименты были проведены на 14 – 21 день после рождения (p14 – p21), паттерны окрашивания были подтверждены у молодых (p7) и зрелых (6 недель) животных. Иммунофлуоресценция, X - gal, сортировка, RNA-seq, SXT, и биохимические эксперименты проводили на 4 – 8 недельных животных. Для создания tetO - LBR - IRES - GFP мыши и культур клеток, описанных в этой статье, использовали процедуры, описанные в расширенном описании экспериментов. Захват ДНК FISH образцов и анализ микроэррей Описаны на схеме рис 1А и более подробно в расширенном описании экспериментов и таблице S1. ДНК FISH, иммуно ДНК FISH ДНК FISH эксперименты проводили по методу, описанному ранее (Lomvardas et al ., 2006) с изменениями, описанными в таблицах S4 и S5 расширенного описания экспериментов. Микроскопия и анализ изображений. Конфокальные изображения были получены на Zeiss LSM 700. Каналы были псевдоподкрашены для логичности и наглядности. Более подробное описание можно найти в расширенном описании экспериментов. SXT (мягкая рентгенография) Нейроны были диссоциированы при использовании растворения в папаине нейробазальный А посредник с HEPES, глутамин и метилцеллюлоза на 30 – 45 минут после чего реакцию останавливали добавлением альбумина. Клетки отмывали, фильтровали, загружали в капилляры и сфотографированы, как описано ранее ( Uchida et al ., 2009). Статистический анализ проводили с использованием программного обеспечения Amira ( Mercury Computer Systems ). Иммуноокрашивание и антитела ИФ проводили в стандартных условиях на МОЕ криосрезах, использованные антитела представлены в Таблице S5. LBR ИФ проводили со стандартными антителами к мышиному LBR (Olins et al ., 2009). См. также в расширенном описании экспериментов. ДНКазный анализ и нативный ChIP ДНКазный анализ и нативный ChIP проводили как описано ранее ( Mag klara et al ., 2011), также см. таблицу S6. 4C 4C проводили, как описано ранее ( Lomvardas et al ., 2006). После инверсии ПЦР, продукты были анализированы для обогащения количественным ПЦР. Анализ экспрессии Для рис 6 B нейроны были отобраны и выделена РНК как описано ранее ( Magklara et al ., 2011). Праймеры для количественного обратно-транскриптазного ПЦР представлены в таблице S 6. Для РНК FISH , RNA - seq , X - gal окрашивания, RT - PCR анализ одной клетки. Материал опубликован 23 февраля 2013 г. |
| |
|
|---|











|
|---|